


Research
Biochemistry and Cell Biology of Lipids
Lipids are central for life and their importance as active players in Cell Biology and Physiology has sparked a high scientific interest in the last years. Lipid involvement in many cellular and organismal aspects has therefore triggered research in various fields.
The fact that the cellular lipid pool consists of several hundred individual lipid species is expression for the multitude of lipid functions and the challenge to understand their individual roles.
Lipid metabolic tracing – studying the dynamics of the lipidome
Why?
Fatty acids are major building blocks of membrane lipids and precursors of many signaling substances. In the form of triglycerides, they are abundant components of our nutrition. Major organs of fatty acid metabolism are gut, liver, muscle and adipose tissue, but all other tissues also have the capacity to use fatty acids either for generation of membrane lipids or of metabolic energy. Our lipidome contains several thousands of lipids, most of which contain fatty acids. Accordingly, the metabolism of fatty acids is extremely complex, and it is amazingly fast – a labeled fatty acid is found in hundreds of different compounds after five minutes of metabolism. Understanding this complexity and its pathological deviations needs experimental tools that offer high sensitivity and time resolution.
How?
Tracing needs a tracer that is as similar as possible to the target molecule but nonetheless reliably distinguishable. Our tracers are alkynes - fatty acids and other lipids that contain a terminal triple bond (Kuerschner & Thiele, 2022). We feed them to biological systems in which they are metabolized, and then we collect and analyze the alkyne-containing (= labeled) lipids. For that, we have developed two technologies:
Mass spectrometric tracing
This is the new approach published in 2019 (Thiele et al. 2019) and includes sample multiplexing capabilities. Extracted alkyne lipids are reacted with the azide reporter C171 or the azide reporters C175 for sample multiplexing.
Importantly, this approach if fully compatible with a parallel use of multiple alkyne lipid tracers as we recently described (Wunderling et al., 2023), offering multiplexed multilabeling analysis of lipid metabolism at unprecedended depth and detail.
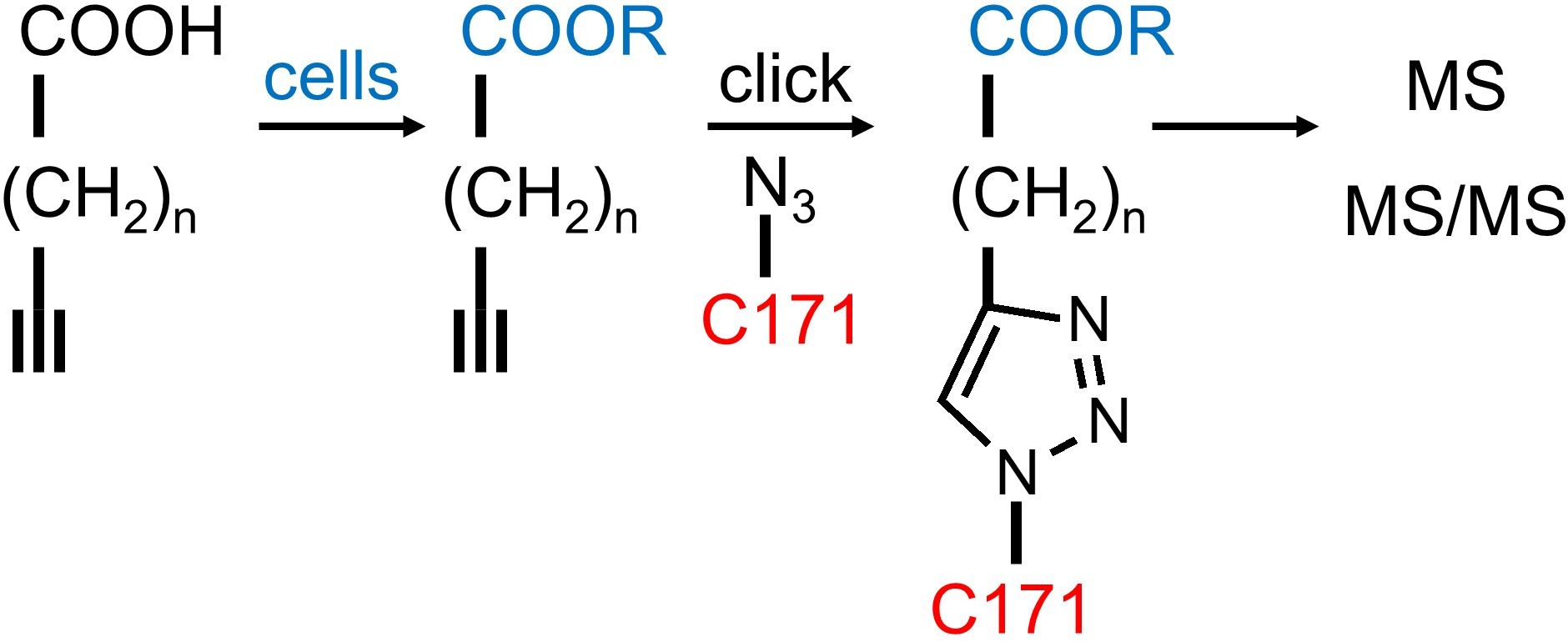
C171 is optimized for improved detection by mass spectrometry. The positive charge strongly improves ionization, and the trialkylammonium group shows predictable fragmentation in tandem MS2.
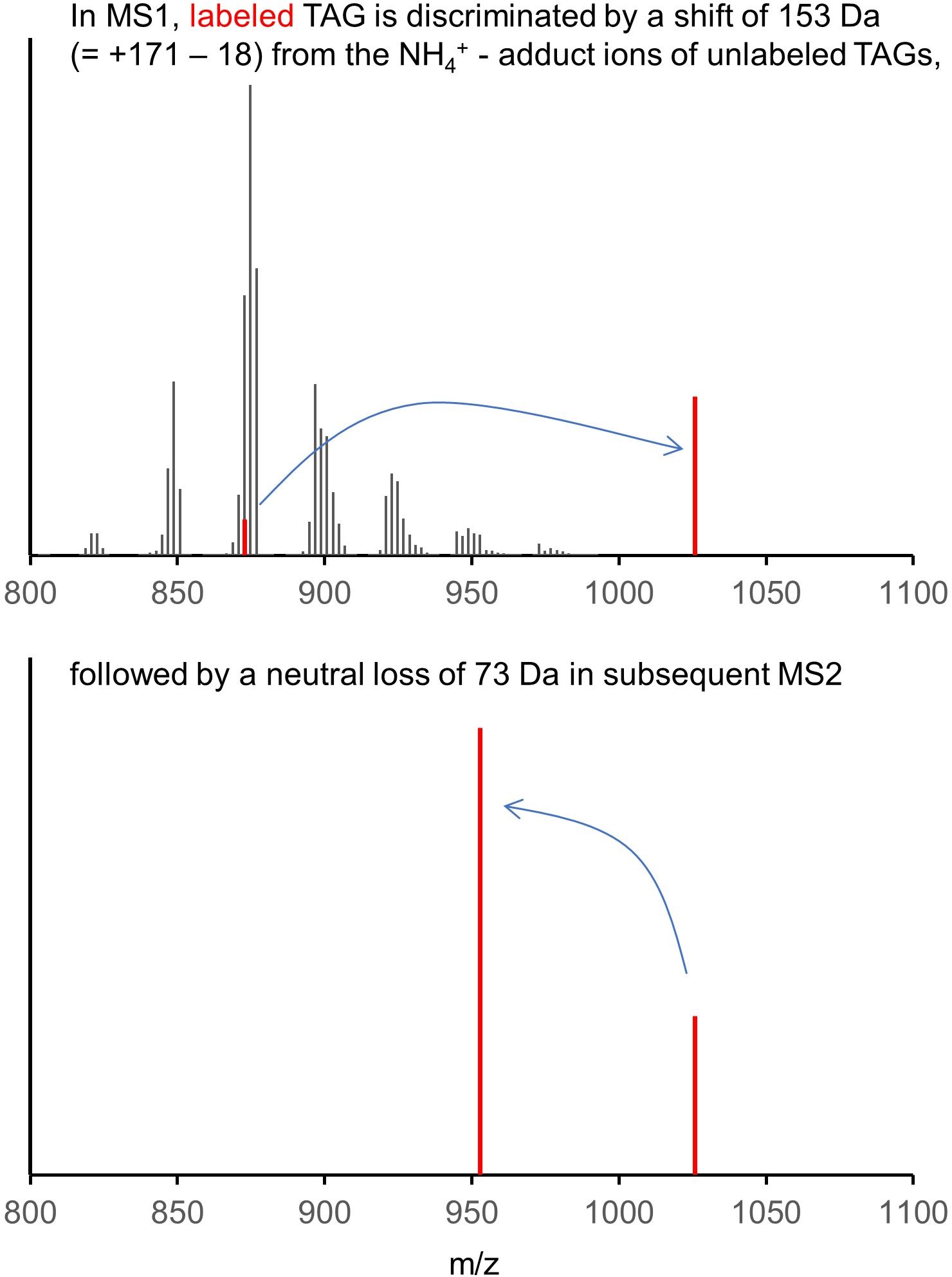
Key advantages are:
- Perfect specificity: labeled species are reliably discriminated from unlabeled species even if present in trace amounts
- Strongly increased sensitivity: improved ionization results in improved sensitivity, typically by 5 – 50-fold.
- Absolut quantification by addition of internal standards
Typical performance (75000 hepatocytes per sample):
- Upon a 5 min labeling, identification of 150 – 250 labeled species
- Upon a 1 h labeling, identification of up to 1000 labeled species
- 15 – 20 lipid classes, 10 with absolute quantification
Single cell analysis:
- Hepatocytes labeled for 3 h
- Single cells by limiting dilution
- 60 – 100 labeled species with absolute quantification
Multiplexing:
A set of deuterated versions of the reagent, the C175 reagents, enables a multiplexed version of the procedure
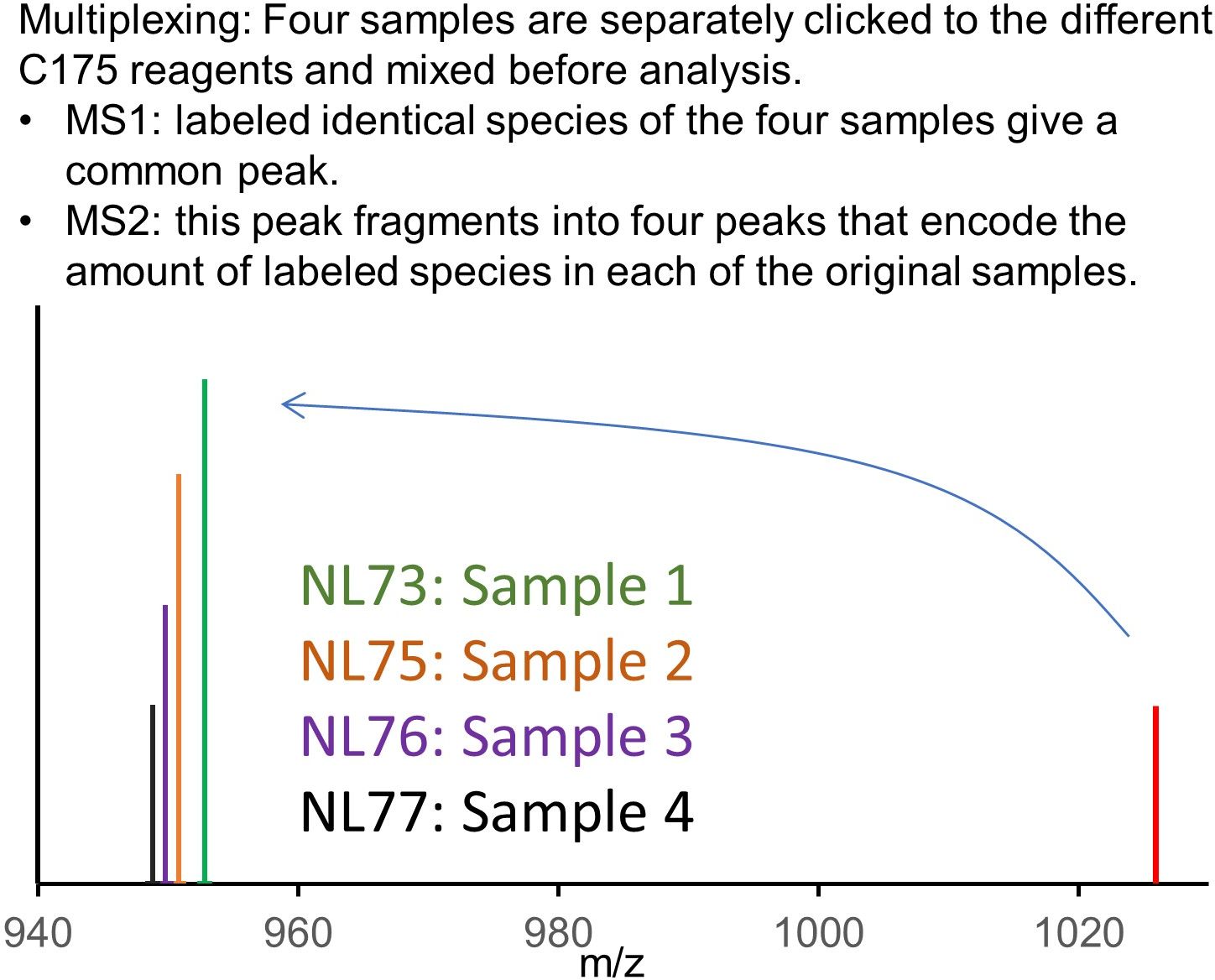
Additional advantages of multiplexing:
- Four fold increased sample turnover
- Improved sample-to-sample comparison by elimination of stochastic variation
Further reading:
Multiplexed and single cell tracing of lipid metabolism: A step by step protocol. Nature Protocol Exchange
And the Behind the Paper blog at Nature’s Protocol and Methods Community
Fluorescent tracing
By reaction with a fluorogenic coumarin dye, labeled lipids become fluorescent. After separation by TLC, the labeled spots can be identified by co-migrating external standards and quantified relative to each other. Absolute quantification is possible but not as reliable as in mass spectrometry.
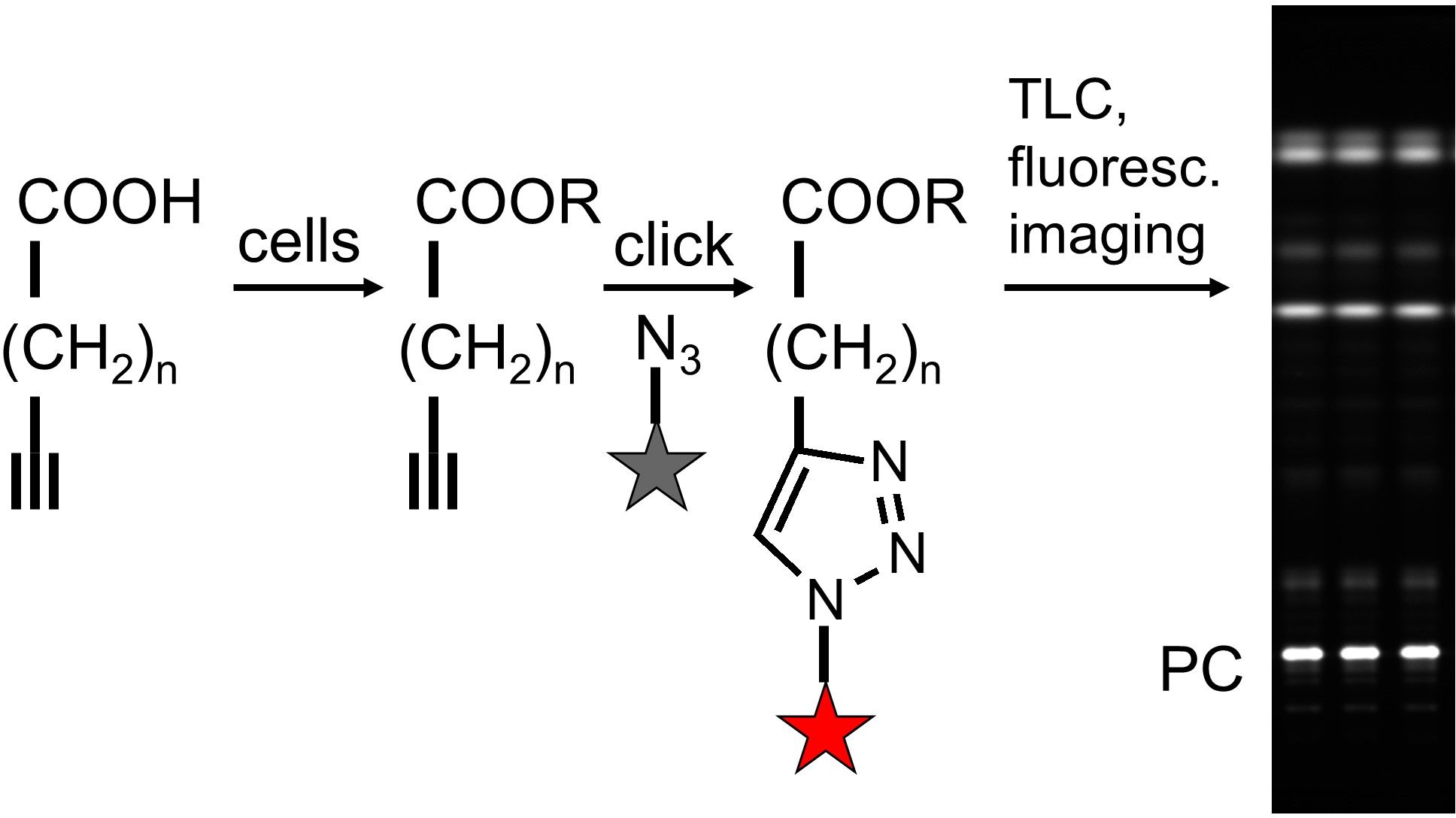
Key advantages:
- Easily accessible
- Parallel sample processing with high throughput
This method was originally published in 2012 (Thiele et al. 2012) and has since then been used in numerous publications in the various fields of lipid research we work in:
Lipid droplets - Function, regulation and composition
- Local lipid metabolism and protein degradation
Lipid droplets (LDs) are the main cellular site for neutral lipid storage but are also important for lipid metabolism, proteasomal protein degradation and autophagy.
We have extensively studied the LD organelle and its lipid and protein content (Spandl et al. 2008), (Grimard et al. 2008). We were the first to demonstrate the presence of the enzymes diacylglycerol-acyltransferase 2 (DGAT2) and lyso-phosphatidyl-acyltransferases 1/2 (LPCAT1/2) at LDs and to show that these enzymes locally synthesize neutral and phospholipids, respectively (Kuerschner et al. 2008), (Moessinger et al. 2011).
It was concluded that LDs have the capacity to adapt their surface to volume ratio by local synthesis of neutral and phospholipids independent of direct connections to the endoplasmic reticulum (Penno et al. 2013), (Moessinger et al. 2014).
We also found the ancient ubiquitous protein 1 (AUP1), a protein involved in proteasomal protein degradation, localizing to LDs in a monotopic fashion. Locally, the highly conserved AUP1 engages in a complex with the Ube2g2 protein and thereby establishes a direct molecular link between the LD organelle and the cellular ubiquitination machinery (Spandl et al. 2011), (Stevanovic et al. 2013), (Lohmann et al. 2013).
We were the first to quantitatively study the rate of triglyceride cycling in adiopcytes (Wunderling et al. 2023). This continuous degradation and re-synthesis we found linked to turnover and re-arrangement of fatty acids with an estimated half life of 2-4 h in these cells. Through cycling and modification, saturated fatty acids are slowly converted to monounsaturated fatty acids, and linoleic acid to arachidonic acid (Thiele et al 2023). The overall process facilitates cellular adjustments to the stored fatty acid pool to meet changing needs of the cell.
Lipid-protein interactions
- Photoactivatable lipid probes
Although all membrane proteins closely interact with lipids, these protein-lipid interactions are only infrequently studied due to technical challenges. Our photoactivatable lipids yielded important new insight into the role of lipids in membrane traffic (Suchanek et al. 2007, Thiele et al. 2000, Schmidt et al. 1999).
Using our 'photocholesterol' we were able to prove the interaction of oxysterol-binding protein related proteins (OPRs) and cholesterol. A detailed picture of the active cholesterol-binding residues was drawn by the combination of cholesterol-photocrosslinking assays and site-directed mutagenesis of ORP2 (Suchanek et al. 2007).
The 'polyene lipids' developed in our lab (Kuerschner et al. 2005) have also been used as fluorescent lipid probes to assess lipid-lipid and lipid-protein interactions by FRET analysis (Ernst et al. 2012), (Contreras et al. 2012).
Lipidomics
Lipidomics and cutting edge lipid detection
Several hundred individual lipid species are found in biological systems illustrating the wealth of their biological functions. Mass spectrometry (MS) is the key analytical technology for lipid identification and quantitation (Papan et al. 2014).
Our laboratory operates the LIMES central MS platform equipped with a Thermo Q Exactive Plus that is mainly used for lipidomics and metabolomics, instrumental for research on lipid metabolism, metabolic diseases and their relation to the immune system. Its precision of typically 0.001 Da allows identification of substances just by their mass, while its resolution of 280,000 allows separation of mass peaks that differ by not more than 0.002 Da.
Apart from shutgun lipidomics analysis our key expertise at the LIMES central MS platform as outlined above is the ultra-sensitive detection of alkyne lipid tracers (Kuerschner & Thiele, 2022).
Of note, these tracers offer multi-modal lipid detection possiblities (Kuerschner & Thiele, 2014) as they are also amenable to highly sensitve thin-layer chromatography (Thiele et al. 2012), (Gaebler et al. 2013) or fluorescence microscopy (Hofmann et al. 2014), (Kuerschner et al. 2014), (Gaebler et al. 2016), (Alecu et al. 2016), (Lauterbach et al. 2021).